Il diidrossiacetone fosfato È un composto chimico abbreviato con l'acronimo DHAP. È un intermediario in alcune vie metaboliche degli organismi viventi, come la degradazione glicolitica o la glicolisi, nonché nel ciclo di Calvin nelle piante.
Biochimicamente, il DHAP è il prodotto dell'azione di un enzima aldolasi sul fruttosio-1,6-bisfosfato (FBP), che causa una degradazione aldolitica risultante in due composti a tre atomi di carbonio: DHAP e gliceraldeide 3-fosfato (GAP).
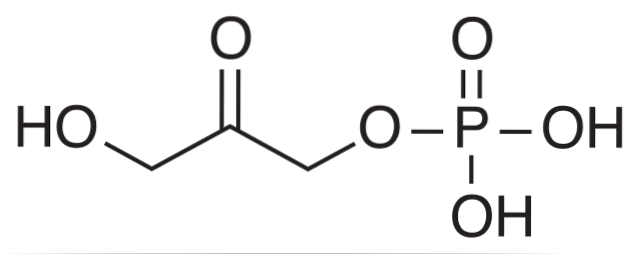
Nel ciclo di Calvin, l'aldolasi esegue la reazione inversa, condensando le molecole di DHAP con le molecole di GAP per formare un esoso..
Indice articolo
Il DHAP è classificato all'interno delle molecole note come chetotriosi. Sono monosaccaridi costituiti da una catena di tre atomi di carbonio (triosi) con il gruppo carbonile sul carbonio centrale (C2).
GAP e DAHP sono isomeri funzionali e costituiscono i carboidrati più semplici all'interno di molecole organiche biologicamente attive.
Sebbene la struttura chimica di molti carboidrati comuni come GAP e DHAP siano aldeidi e chetoni, viene dato loro il termine carboidrato, in riferimento ai derivati diretti dei saccaridi.
Nella glicolisi una serie di reazioni degradano il glucosio in piruvato. Questa degradazione avviene progressivamente in 10 fasi consecutive, in cui intervengono diversi enzimi e si producono una varietà di intermedi, tutti fosforilati..
Il DHAP appare nella glicolisi nella quarta reazione di questo processo, che consiste nella scomposizione di FBP in due carboidrati di tre atomi di carbonio (triosi), di cui solo GAP continua la sequenza della glicolisi, mentre DHAP deve essere trasformato in GAP per seguire questa rotta.
Questa reazione è catalizzata da una aldolasi (fruttosio bisfosfato aldolasi), che effettua una scissione aldolica tra i carboni C3 e C4 del FBP.
Questa reazione si verifica solo se l'esosio da dividere ha un gruppo carbonile in C2 e un idrossile in C4. Per questo motivo, l'isomerizzazione del glucosio-6-fosfato (G6P) in fruttosio-6-fosfato (F6P) si verifica in precedenza..
Il DHAP è anche coinvolto nella quinta reazione di glicolisi, essendo la sua isomerizzazione a GAP da parte dell'enzima trioso fosfato isomerasi o TIM. Con questa reazione si completa la prima fase di degradazione del glucosio..
Nella ripartizione aldolica si producono due intermedi, dove DHAP costituisce il 90% della miscela in equilibrio.
Esistono due tipi di aldolasi: a) l'aldolasi di tipo I è presente nelle cellule animali e vegetali ed è caratterizzata dalla formazione di una base di Schiff tra il sito attivo enzimatico e il carbonile della FBP. b) L'aldolasi di tipo II si trova in alcuni batteri e funghi, ha un metallo nel sito attivo (solitamente Zn).
La scissione aldolica inizia con l'adesione del substrato al sito attivo e la rimozione di un protone dal gruppo β-idrossile, formando la base di Schiff protonata (catione imminio). La scomposizione dei carboni C3 e C4 produce il rilascio di GAP e la formazione di un intermedio chiamato enammina..
L'enammina viene successivamente stabilizzata, si forma così un catione imminio che viene idrolizzato, con il quale viene finalmente rilasciato DHAP e si rigenera così l'enzima libero.
Nelle cellule con aldolasi di tipo II, la formazione della base di Schiff non si verifica, essendo un catione metallico bivalente, generalmente ZnDue+, quello che stabilizza l'enammina intermedia per il rilascio di DHAP.
Come accennato, la concentrazione di equilibrio di DHAP è superiore a quella di GAP, quindi le molecole di DHAP vengono trasformate in GAP poiché quest'ultimo viene utilizzato nella successiva reazione di glicolisi..
Questa trasformazione avviene grazie all'enzima TIM. Questa è la quinta reazione del processo di degradazione glicolitica e in essa i carboni C1 e C6 del glucosio diventano i carboni C3 di GAP, mentre i carboni C2 e C5 diventano C2 e C3 e C4 del glucosio diventano C1 del GAP.
L'enzima TIM è considerato l '"enzima perfetto" perché la diffusione controlla la velocità della reazione, il che significa che il prodotto si forma con la stessa rapidità con cui il sito attivo dell'enzima e il suo substrato si uniscono..
Nella reazione di trasformazione del DHAP in GAP si forma un intermedio chiamato enediol. Questo composto è in grado di cedere i protoni dei gruppi idrossilici a un residuo del sito attivo dell'enzima TIM..
Il ciclo di Calvin è il ciclo di riduzione fotosintetica del carbonio (PCR) che costituisce la fase oscura del processo di fotosintesi nelle piante. In questa fase, i prodotti (ATP e NADPH) ottenuti nella fase leggera del processo vengono utilizzati per produrre carboidrati..
In questo ciclo si formano sei molecole GAP, di cui due vengono trasformate in DHAP per isomerizzazione, grazie all'azione dell'enzima TIM, in reazione inversa a quella che avviene nella degradazione della glicolisi. Questa reazione è reversibile, sebbene l'equilibrio, nel caso di questo ciclo e, a differenza della glicolisi, sia spostato verso la conversione di GAP in DHAP.
Queste molecole DHAP possono quindi seguire due percorsi, uno è una condensazione aldolica catalizzata da un'aldolasi, in cui si condensa con una molecola GAP per formare FBP.
L'altra reazione che uno dei DHAP può assumere è un'idrolisi del fosfato catalizzata da una sedoeptulosio bisfosfatasi. In quest'ultima via reagisce con un eritrosio formando sedoeptulosio 1,7-bisfosfato.
Nella gluconeogenesi alcuni composti non glucidici come piruvato, lattato e alcuni amminoacidi vengono convertiti in glucosio. In questo processo il DHAP si ripresenta per isomerizzazione di una molecola GAP per azione di TIM, quindi per condensazione aldolica diventa FBP.



Nessun utente ha ancora commentato questo articolo.