Il Scatola TATA, Nella biologia cellulare, è una sequenza di DNA di consenso che si trova in tutte le linee di organismi viventi ed è ampiamente conservata. La sequenza è 5'-TATAAA-3 'e può essere seguita da alcune adenine ripetute.
La posizione della scatola è a monte (o a monte, come viene spesso chiamata in letteratura) dall'inizio della trascrizione. Questo si trova nella regione del promotore dei geni, dove si verificherà l'unione con i fattori di trascrizione. Oltre a questi fattori, l'RNA polimerasi II tende a legarsi alla scatola TATA..
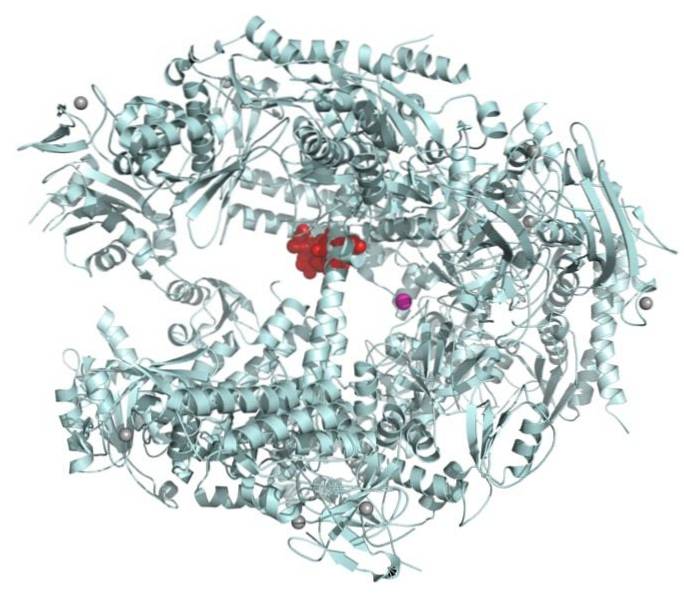
Sebbene la scatola TATA sia la sequenza principale del promotore, ci sono geni che ne sono privi.
Indice articolo
L'inizio della sintesi dell'RNA richiede che l'RNA polimerasi si leghi a sequenze specifiche nel DNA, chiamate promotori. La casella TATA è la sequenza di consenso di un promotore. È chiamata la scatola di Pribnow nei procarioti e la scatola di Goldberg-Hogness negli eucarioti.
Pertanto, la scatola TATA è una regione conservata nel DNA. Il sequenziamento di numerose regioni di inizio trascrizione del DNA ha dimostrato che la sequenza di consenso, o sequenza comune, è (5ʾ) T * A * TAAT * (3ʾ). Le posizioni contrassegnate da un asterisco hanno un'omologia elevata. L'ultimo residuo T si trova sempre nei promotori di E. coli.
Per convenzione, alle coppie di basi che corrispondono all'inizio della sintesi di una molecola di RNA vengono assegnati numeri positivi e alle coppie di basi che precedono l'inizio dell'RNA vengono assegnati numeri negativi. La scatola TATA è nella regione -10.
Sopra E. coli, la regione del promotore è compresa tra le posizioni -70 e +30. In questa regione c'è una seconda sequenza di consenso (5ʾ) T * TG * ACA (3ʾ) alla posizione -35. Allo stesso modo, le posizioni contrassegnate da un asterisco hanno un'omologia elevata..
Negli eucarioti, le regioni del promotore hanno elementi di segnale che differiscono per ciascuna delle RNA polimerasi. Sopra E. coli una singola RNA polimerasi identifica gli elementi segnale nella regione del promotore.
Inoltre, negli eucarioti le regioni promotrici sono più diffuse. Esistono diverse sequenze, situate nella regione -30 e -100, che stabiliscono diverse combinazioni nei diversi promotori.
Negli eucarioti, ci sono numerosi fattori di trascrizione che interagiscono con i promotori. Ad esempio, il fattore TFIID si lega alla sequenza TATA. D'altra parte, i geni dell'RNA ribosomiale sono strutturati sotto forma di più geni, uno seguito dall'altro..
Le variazioni nelle sequenze di consenso delle regioni -10 e -35 alterano il legame della RNA polimerasi alla regione del promotore. Pertanto, una singola mutazione della coppia di basi causa una diminuzione della velocità di legame della RNA polimerasi alla regione del promotore..
La scatola TATA partecipa al legame e all'inizio della trascrizione. Sopra E. coli, l'oloenzima della RNA polimerasi è costituito da cinque subunità αDueββσ. La subunità σ si lega al DNA a doppia elica e si sposta alla ricerca del box TATA, che è il segnale che indica l'inizio del gene.
La subunità σ della RNA polimerasi ha una costante di associazione del promotore molto alta (nell'ordine 10undici), che indica un'elevata specificità di riconoscimento tra esso e la sequenza di box di Pribnow.
La RNA polimerasi si lega al promotore e forma un complesso chiuso. Quindi forma un complesso aperto caratterizzato dall'apertura locale di 10 paia di basi della doppia elica del DNA. Questa apertura è facilitata perché la sequenza del box Pribnow è ricca di A-T.
Quando il DNA viene svolto, si forma il primo legame fosfodiestere e inizia l'allungamento dell'RNA. La subunità σ viene rilasciata e la RNA polimerasi lascia il promotore. Altre molecole di RNA polimerasi possono legarsi al promotore e iniziare la trascrizione. In questo modo, un gene può essere trascritto molte volte..
Nel lievito, l'RNA polimerasi II è costituita da 12 subunità. Questo enzima avvia la trascrizione riconoscendo due tipi di sequenze di consenso all'estremità 5ʾ dell'inizio della trascrizione, vale a dire: sequenza di consenso TATA; Sequenza di consenso CAAT.
La RNA polimerasi II richiede proteine, chiamate fattori di trascrizione TFII, per formare un complesso di trascrizione attivo. Questi fattori sono abbastanza conservati in tutti gli eucarioti..
I fattori di trascrizione sono molecole proteiche che possono legarsi alla molecola di DNA e hanno la capacità di aumentare, diminuire o arrestare la produzione di un gene specifico. Questo evento è cruciale per la regolazione genica.
La formazione del complesso di trascrizione inizia con il legame della proteina TBP ("proteina legante TATA") alla scatola TATA. A sua volta, questa proteina lega TFIIB, che si lega anche al DNA. Il complesso TBP-TFIIB si lega a un altro complesso costituito da TFIIF e RNA polimerasi II. In questo modo, TFIIF aiuta la RNA polimerasi II a legarsi al promotore..
Alla fine, TFIIE e TFIIH si uniscono e creano un complesso chiuso. TFIIH è un'elicasi e promuove la separazione del doppio filamento del DNA, un processo che richiede ATP. Ciò accade vicino al sito di inizio della sintesi dell'RNA. In questo modo si forma il complesso aperto.
La proteina p53 è un fattore di trascrizione, noto anche come proteina soppressore del tumore p53. È il prodotto di un oncogene dominante. La sindrome di Li-Fraumeni è causata da una copia di questo gene mutato, che causa lo sviluppo di carcinomi, leucemie e tumori.
È noto che P53 inibisce la trascrizione di alcuni geni e attiva quella di altri. Ad esempio, p53 impedisce la trascrizione dei geni con un promotore TATA formando un complesso costituito da p53, altri fattori di trascrizione e il promotore TATA. Pertanto, p53 mantiene la crescita cellulare sotto controllo..

Nessun utente ha ancora commentato questo articolo.