Il topoisomerasi Sono un tipo di enzimi isomerasi che modificano la topologia dell'acido desossiribonucleico (DNA), generando sia il suo svolgimento che il suo avvolgimento e superavvolgimento.
Questi enzimi hanno un ruolo specifico nell'alleviare lo stress torsionale nel DNA in modo che possano verificarsi processi importanti come la replicazione, la trascrizione del DNA nell'acido ribonucleico messaggero (mRNA) e la ricombinazione del DNA..
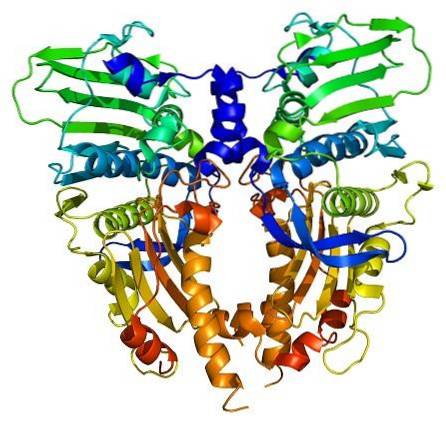
Gli enzimi topoisomerasi sono presenti sia nelle cellule eucariotiche che in quelle procariotiche. La sua esistenza è stata prevista dagli scienziati Watson e Crick, nel valutare i limiti che la struttura del DNA presentava per consentire l'accesso alle sue informazioni (memorizzate nella sua sequenza nucleotidica).
Per comprendere le funzioni delle topoisomerasi, si deve considerare che il DNA ha una struttura a doppia elica stabile, con i suoi fili avvolti l'uno sull'altro..
Queste catene lineari sono costituite da 2-desossiribosio legato da legami fosfodiestere 5'-3 'e basi azotate al loro interno, come i pioli di una scala a chiocciola..
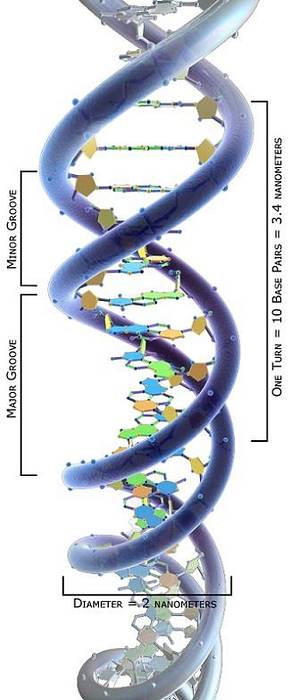
Lo studio topologico delle molecole di DNA ha dimostrato che possono assumere varie conformazioni dipendenti dal loro stress torsionale: da uno stato rilassato a diversi stati di avvolgimento che ne consentono la compattazione..
Le molecole di DNA con diverse conformazioni sono chiamate topoisomeri. Quindi, possiamo concludere che le topoisomerasi I e II possono aumentare o diminuire lo stress torsionale delle molecole di DNA, formando i loro diversi topoisomeri..
Tra i possibili topoisomeri a DNA, la conformazione più comune è il supercoil, che è molto compatto. Tuttavia, la doppia elica del DNA deve essere svolta anche dalle topoisomerasi durante vari processi molecolari..
Indice articolo
Alcune topoisomerasi possono rilassare solo le supercoil negative del DNA, o entrambe le supercoil del DNA: positiva e negativa.
Se il DNA circolare a doppia elica viene svolto sul suo asse longitudinale e si verifica una rotazione sinistrorsa (in senso orario), si dice che sia superavvolto negativamente. Se il giro è in senso orario (senso antiorario), è positivamente avvolto.

Fondamentalmente, le topoisomerasi possono:
-Facilitare il passaggio di un filamento di DNA attraverso un taglio nel filamento opposto (topoisomerasi di tipo I).
-Facilitare il passaggio di una doppia elica completa attraverso una scissione stessa, o attraverso una scissione in una doppia elica diversa (topoisomerasi di tipo II).
In sintesi, le topoisomerasi agiscono attraverso la scissione dei legami fosfodiestere, in uno o entrambi i filamenti che compongono il DNA. Quindi modificano lo stato di avvolgimento dei fili di una doppia elica (topoisomerasi I) o di due doppie eliche (topoisomerasi II), per legare o legare di nuovo le estremità tagliate.
Sebbene la topoisomerasi I sia un enzima che mostra una maggiore attività durante la fase S (sintesi del DNA), non si ritiene che dipenda da una fase del ciclo cellulare.
Mentre l'attività della topoisomerasi II è più attiva durante la fase logaritmica della crescita cellulare e nelle cellule di tumori a crescita rapida.
L'alterazione dei geni che codificano per la topoisomerasi è letale per le cellule, evidenziando l'importanza di questi enzimi. Tra i processi a cui partecipano le topoisomerasi ci sono:
Le topoisomerasi facilitano l'archiviazione delle informazioni genetiche in modo compatto, poiché generano l'avvolgimento e il superavvolgimento del DNA, consentendo di trovare una grande quantità di informazioni in un volume relativamente piccolo.
Senza le topoisomerasi e le loro caratteristiche uniche, l'accesso alle informazioni immagazzinate nel DNA sarebbe impossibile. Ciò è dovuto al fatto che le topoisomerasi rilasciano periodicamente lo stress torsionale che si genera nella doppia elica del DNA, durante il suo svolgimento, nei processi di replicazione, trascrizione e ricombinazione..
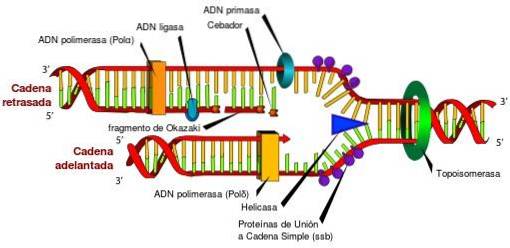
Se lo stress torsionale generato durante questi processi non viene rilasciato, potrebbe verificarsi un'espressione genica difettosa, l'interruzione del DNA o cromosoma circolare, producendo anche la morte cellulare.
I cambiamenti conformazionali (nella struttura tridimensionale) della molecola di DNA espongono all'esterno regioni specifiche, che possono interagire con le proteine leganti il DNA. Queste proteine hanno una funzione di regolazione dell'espressione genica (positiva o negativa).
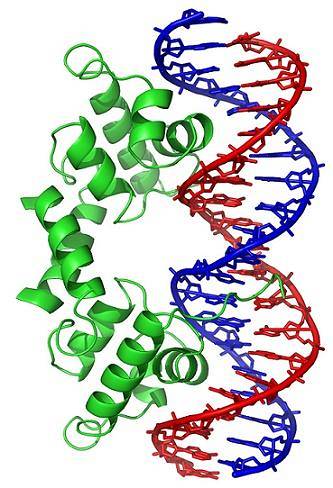
Pertanto, lo stato di avvolgimento del DNA, generato dall'azione delle topoisomerasi, influisce sulla regolazione dell'espressione genica.
La topoisomerasi II è necessaria per l'assemblaggio dei cromatidi, la condensazione e la decondensazione dei cromosomi e la segregazione delle molecole di DNA figlie durante la mitosi..
Questo enzima è anche una proteina strutturale e uno dei principali costituenti della matrice del nucleo cellulare durante l'interfase..
Esistono due tipi principali di topoisomerasi a seconda che siano in grado di scindere uno o due filamenti di DNA.
Le topoisomerasi di tipo I sono monomeri che alleviano i supercoil negativi e positivi, prodotti dal movimento delle forcine durante la trascrizione e durante i processi di replicazione e ricombinazione genica..
Le topoisomerasi di tipo I possono essere suddivise in tipo 1A e tipo 1B. Questi ultimi sono quelli che si trovano negli esseri umani e sono responsabili del rilassamento del DNA superavvolto.
La topoisomerasi 1B (Top1B) è composta da 765 aminoacidi suddivisi in 4 domini specifici. Uno di questi domini ha un'area altamente conservata contenente il sito attivo della tirosina (Tyr7233). Tutte le topoisomerasi presentano nel loro sito attivo una tirosina con un ruolo fondamentale nell'intero processo catalitico.
La tirosina del sito attivo forma un legame covalente con l'estremità 3'-fosfato del filamento di DNA, tagliandolo e mantenendolo attaccato all'enzima, mentre fa passare un altro filamento di DNA attraverso la scissione.
Il passaggio dell'altro filamento di DNA attraverso il filamento diviso è ottenuto grazie ad una trasformazione conformazionale dell'enzima, che produce l'apertura della doppia elica del DNA..
Quindi la topoisomerasi I ritorna alla sua conformazione iniziale e lega di nuovo le estremità tagliate. Ciò avviene tramite un processo inverso alla rottura della catena del DNA, nel sito catalitico dell'enzima. Infine, la topoisomerasi rilascia il filamento di DNA.
Il tasso di legatura del DNA è superiore al tasso di escissione, garantendo così la stabilità della molecola e l'integrità del genoma..
In sintesi, la topoisomerasi di tipo I catalizza:
Le topoisomerasi di tipo II sono enzimi dimerici, che scindono entrambi i filamenti di DNA, rilassando così i supercoil generati durante la trascrizione e altri processi cellulari.
Questi enzimi hanno bisogno di magnesio (Mg++) e necessitano anche dell'energia che proviene dalla rottura del legame trifosfato ATP, di cui sfruttano grazie ad un ATPasi.
Le topoisomerasi II umane sono molto simili a quelle del lievito (Saccharomyces cerevisiae), che si compone di due monomeri (sottoframmenti A e B). Ogni monomero ha un dominio ATPasi e in un sotto-frammento il sito attivo tirosina 782, a cui il DNA può legarsi. Pertanto, due filamenti di DNA possono legarsi alla topoisomerasi II..
Il meccanismo d'azione della topoisomerasi II è lo stesso descritto per la topoisomerasi I, considerando che due filamenti di DNA sono separati e non solo uno..
Nel sito attivo della topoisomerasi II, un frammento di doppia elica di DNA, chiamato "frammento G". Questo frammento viene asportato e tenuto insieme al sito attivo da legami covalenti..
Quindi, l'enzima fa passare un altro frammento di DNA, chiamato "frammento T", attraverso il frammento scisso "G", grazie a un cambiamento conformazionale dell'enzima, che dipende dall'idrolisi dell'ATP.
La topoisomerasi II lega le due estremità del "frammento G" e alla fine recupera il suo stato iniziale, rilasciando il frammento "G". Il DNA quindi rilassa lo stress torsionale, consentendo il verificarsi dei processi di replicazione e trascrizione..
Il genoma umano ha cinque topoisomerasi: top1, top3α, top3β (di tipo I); e top2α, top2β (di tipo II). Le topoisomerasi umane più rilevanti sono top1 (topoisomerasi di tipo IB) e 2α (topoisomerasi di tipo II).
Poiché i processi catalizzati dalle topoisomerasi sono necessari per la sopravvivenza delle cellule, questi enzimi sono un buon bersaglio di attacco per colpire le cellule maligne. Per questo motivo, le topoisomerasi sono considerate importanti nel trattamento di molte malattie umane..
I farmaci che interagiscono con le topoisomerasi sono attualmente ampiamente studiati come sostanze chemioterapiche contro le cellule tumorali (in diversi organi del corpo) e microrganismi patogeni..
I farmaci che inibiscono l'attività della topoisomerasi possono:
La stabilizzazione del complesso transitorio che si forma dal legame del DNA alla tirosina del sito catalitico dell'enzima, impedisce il legame dei frammenti scissi, che può portare alla morte cellulare.
I composti che inibiscono la topoisomerasi includono i seguenti.
Gli antibiotici sono usati contro il cancro, poiché impediscono la crescita delle cellule tumorali, di solito interferendo con il loro DNA. Questi sono spesso chiamati antibiotici antineoplastici (cancro). L'actinomicina D, ad esempio, colpisce la topoisomerasi II ed è utilizzata nei tumori di Wilms nei bambini e nei rabdomiosarcomi.
Le antracicline sono, tra gli antibiotici, uno dei farmaci antitumorali più efficaci e con lo spettro più ampio. Sono usati per trattare il cancro del polmone, delle ovaie, dell'utero, dello stomaco, della vescica, della mammella, della leucemia e dei linfomi. È noto che influisce sulla topoisomerasi II per intercalazione nel DNA.
La prima antraciclina isolata da un actinobacteria (Streptomyces peucetius) era la daunorubicina. Successivamente la doxorubicina è stata sintetizzata in laboratorio e attualmente vengono utilizzate anche epirubicina e idarubicina.
Gli antracinoni o antracenedioni sono composti derivati dall'antracene, simili alle antracicline, che influenzano l'attività della topoisomerasi II per intercalazione nel DNA. Sono usati per il cancro al seno metastatico, il linfoma non Hodgkin (NHL) e la leucemia..
Questi farmaci sono stati trovati nei pigmenti di alcuni insetti, piante (frangula, senna, rabarbaro), licheni e funghi; così come hoelite, che è un minerale naturale. A seconda della dose, possono essere cancerogeni.
Tra questi composti, abbiamo il mitoxantrone e il suo analogo, losoxantrone. Questi impediscono la proliferazione di cellule tumorali maligne, legandosi al DNA in modo irreversibile.
Le podofillotossine, come le epidofillotossine (VP-16) e il teniposide (VM-26), formano un complesso con la topoisomerasi II. Sono usati contro il cancro ai polmoni, ai testicoli, alla leucemia, ai linfomi, al cancro dell'ovaio, al carcinoma mammario e ai tumori intracranici maligni, tra gli altri. Sono isolati dalle piante Podophyllum notatum Y P. peltatum.
Le campotecine sono composti che inibiscono la topoisomerasi I, inclusi irinotecan, topotecan e diflomotecan..
Questi composti sono stati usati contro il cancro del colon, del polmone e della mammella e sono ottenuti naturalmente dalla corteccia e dalle foglie delle specie arboree. Camptotheca acuminata delle cornee cinesi e del Tibet.
Anche le alterazioni strutturali delle topoisomerasi I e II possono verificarsi in modo completamente naturale. Questo può accadere durante alcuni eventi che influenzano il tuo processo catalitico.
Queste alterazioni includono la formazione di dimeri di pirimidina, disadattamento di basi di azoto e altri eventi causati dallo stress ossidativo..



Nessun utente ha ancora commentato questo articolo.