Il nucleosoma è l'unità di base del confezionamento del DNA negli organismi eucarioti. È quindi l'elemento di compressione della cromatina più piccolo.
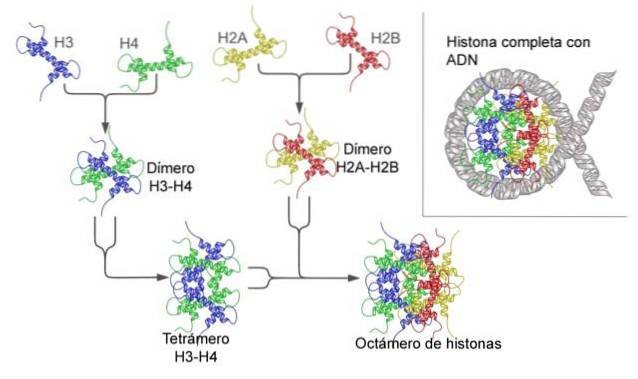
Il nucleosoma è costruito come un ottamero di proteine chiamate istoni, o una struttura a forma di tamburo su cui sono avvolti circa 140 nt di DNA, compiendo quasi due giri completi..
Inoltre, un ulteriore 40-80 nt di DNA è considerato parte del nucleosoma ed è la frazione di DNA che consente la continuità fisica tra un nucleosoma e l'altro in strutture di cromatina più complesse (come la fibra di cromatina da 30 nm)..
Il codice istonico è stato uno dei primi elementi di controllo epigenetico compreso a livello molecolare..
Indice articolo
I nucleosomi consentono:
Nella sua accezione più elementare, i nucleosomi sono costituiti da DNA e proteine. Il DNA può essere virtualmente qualsiasi DNA a doppia banda presente nel nucleo della cellula eucariotica, mentre le proteine nucleosomiche appartengono tutte all'insieme di proteine chiamate istoni..
Gli istoni sono piccole proteine con un alto carico di residui amminoacidici basici; Ciò consente di contrastare l'elevata carica negativa del DNA e stabilire un'efficace interazione fisica tra le due molecole senza raggiungere la rigidità del legame chimico covalente..
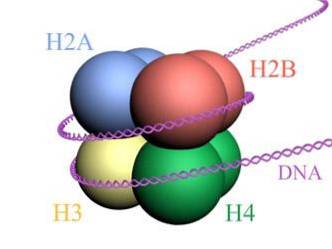
Gli istoni formano un ottamero simile a un tamburo con due copie o monomeri di ciascuno degli istoni H2A, H2B, H3 e H4. Il DNA fa quasi due giri completi sui lati dell'ottamero e poi continua con una frazione di DNA linker che si associa all'istone H1, per tornare a dare due giri completi su un altro istone ottamero..
Il set di ottameri, il DNA associato e il suo corrispondente DNA linker, è un nucleosoma.
Il DNA genomico è costituito da molecole estremamente lunghe (più di un metro nel caso dell'uomo, considerando tutti i loro cromosomi), che devono essere compattate e organizzate all'interno di un nucleo estremamente piccolo.
Il primo passo in questa compattazione viene effettuato attraverso la formazione di nucleosomi. Con questo solo passaggio, il DNA viene compattato circa 75 volte.
Ciò dà origine a una fibra lineare da cui vengono costruiti i livelli successivi di compattazione della cromatina: la fibra da 30 nm, i loop e i loop dei loop..
Quando una cellula si divide, sia per mitosi che per meiosi, il massimo grado di compattazione è rispettivamente il cromosoma mitotico o meiotico stesso..
Il fatto che gli ottameri istonici e il DNA interagiscano elettrostaticamente spiega in parte la loro effettiva associazione, senza perdere la fluidità necessaria per rendere i nucleosomi elementi dinamici per compattare e decompattare la cromatina..
Ma c'è un elemento di interazione ancora più sorprendente: le estremità N-terminali degli istoni sono esposte all'esterno dell'interno dell'ottamero, che è più compatto e inerte..
Queste estremità non solo interagiscono fisicamente con il DNA, ma subiscono anche una serie di modificazioni covalenti dalle quali dipenderà il grado di compattazione della cromatina e l'espressione del DNA associato..
L'insieme di modifiche covalenti, in termini di tipo e numero, tra le altre cose, è noto collettivamente come codice istonico. Queste modifiche includono fosforilazione, metilazione, acetilazione, ubiquitinazione e sumoilazione di residui di arginina e lisina ai terminali N degli istoni..
Ogni cambiamento, insieme ad altri all'interno della stessa molecola o in residui di altri istoni, in particolare istoni H3, determinerà l'espressione o meno del DNA associato, nonché il grado di compattazione della cromatina..
Come regola generale, si è visto, ad esempio, che gli istoni ipermetilati e ipoacetilati determinano che il DNA associato non è espresso e che la cromatina è presente in uno stato più compatto (eterocromatico, e quindi inattivo)..
Al contrario, il DNA eucromatico (meno compatto e geneticamente attivo) è associato a una cromatina i cui istoni sono iperacetilati e ipometilati..
Abbiamo già visto che lo stato di modificazione covalente degli istoni può determinare il grado di espressione e la compattazione locale della cromatina. A livello globale, la compattazione della cromatina è anche regolata da modificazioni covalenti degli istoni nei nucleosomi.
È stato dimostrato, ad esempio, che l'eterocromatina costitutiva (che non è mai espressa ed è densamente impaccata) tende ad aderire alla lamina nucleare, lasciando liberi i pori nucleari..
Da parte sua, l'eucromatina costitutiva (che è sempre espressa, come quella che include i geni del mantenimento cellulare, e si trova nelle regioni della cromatina lassa), lo fa in grandi anelli che espongono il DNA da trascrivere al meccanismo di trascrizione.
Altre regioni del DNA genomico oscillano tra questi due stati a seconda del tempo di sviluppo dell'organismo, delle condizioni di crescita, dell'identità cellulare, ecc..
Per realizzare il loro piano di sviluppo, espressione e mantenimento delle cellule, i genomi degli organismi eucarioti devono regolare con precisione quando e come devono manifestarsi le loro potenzialità genetiche..
A partire quindi dalle informazioni immagazzinate nei loro geni, questi si trovano nel nucleo in particolari regioni che determinano il loro stato trascrizionale..
Possiamo dire, quindi, che un altro dei ruoli fondamentali dei nucleosomi, attraverso i cambiamenti della cromatina che aiuta a definire, è l'organizzazione o l'architettura del nucleo che li ospita..
Questa architettura è ereditata ed è filogeneticamente conservata grazie all'esistenza di questi elementi modulari del packaging informativo.


Nessun utente ha ancora commentato questo articolo.