Con "eredità non mendeliana"Si intende qualsiasi modello di eredità in cui i caratteri ereditati non si separano in conformità con le disposizioni delle leggi di Mendel..
Nel 1865 Gregor Mendel, considerato il "padre della genetica", eseguì una serie di incroci sperimentali con piante di pisello, i cui risultati lo portarono a proporre alcuni postulati (leggi di Mendel) che cercavano di dare una spiegazione logica all'eredità. i personaggi tra genitori e figli.
Questo astuto monaco austriaco osservò attentamente la segregazione dei geni parentali e la loro comparsa nella prole come caratteri dominanti e recessivi. Inoltre, ha determinato i modelli matematici che descrivevano l'ereditarietà da una generazione all'altra e questi risultati sono stati "ordinati" sotto forma di 3 leggi fondamentali:
- La legge del dominio
- La legge della segregazione dei personaggi e
- La legge della distribuzione indipendente.
I successi e le deduzioni di Mendel sono stati nascosti per molti anni, fino alla loro riscoperta all'inizio del XX secolo..

A quel tempo, tuttavia, la comunità scientifica manteneva una posizione alquanto scettica riguardo a queste leggi, poiché non sembravano spiegare i modelli di ereditabilità in nessuna specie animale o vegetale, specialmente in quei caratteri determinati da più di un locus..
Per questo motivo, i primi genetisti classificarono i pattern ereditari osservati come "mendeliani" (quelli che potevano essere spiegati attraverso la segregazione di alleli semplici, dominanti o recessivi, appartenenti allo stesso locus) e "non mendeliani" (quelli che non potrebbe essere spiegato così facilmente).
Indice articolo
L'eredità mendeliana si riferisce a un modello ereditario conforme alle leggi di segregazione e distribuzione indipendente, secondo cui un gene ereditato da qualsiasi genitore secerne nei gameti con una frequenza equivalente o, piuttosto, con la stessa probabilità.
I principali modelli di ereditarietà mendeliana che sono stati descritti per alcune malattie sono: autosomica recessiva, autosomica dominante e legata al cromosoma X, che si aggiungono ai modelli di dominanza e recessività descritti da Mendel.
Tuttavia, questi sono stati postulati per quanto riguarda i tratti visibili e non i geni (va notato che alcuni alleli possono codificare tratti che si segregano come dominanti, mentre altri possono codificare gli stessi tratti, ma questi si separano come geni recessivi).
Da quanto sopra, ne consegue che l'eredità non mendeliana consiste, semplicemente, in qualsiasi modello ereditario che non soddisfa la norma in cui un gene ereditato da qualsiasi genitore secerne nelle cellule germinali con le stesse probabilità, e questi includono:
- Eredità mitocondriale
- "Stampa"
- Disomia uniparentale
- Dominanza incompleta
- Codominanza
- Alleli multipli
- Pleiotropia
- Alleli letali
- Tratti poligenici
- Eredità legata al sesso
Il verificarsi di queste variazioni nei modelli ereditari può essere attribuito alle varie interazioni che i geni hanno con altri componenti cellulari, oltre al fatto che ognuno è soggetto a regolazione e variazione in uno qualsiasi degli stadi di trascrizione, splicing, traduzione, proteina piegatura, oligomerizzazione, traslocazione e compartimentazione all'interno della cellula e per la sua esportazione.
In altre parole, ci sono numerose influenze epigenetiche che possono modificare i modelli di ereditarietà di qualsiasi tratto, risultando in una "deviazione" dalle leggi di Mendel..
Il DNA mitocondriale trasmette anche informazioni da una generazione all'altra, proprio come fa ciò che è contenuto nel nucleo di tutte le cellule eucariotiche. Il genoma codificato in questo DNA comprende i geni necessari per la sintesi di 13 polipeptidi che fanno parte delle subunità della catena respiratoria mitocondriale, essenziali per organismi con metabolismo aerobico..
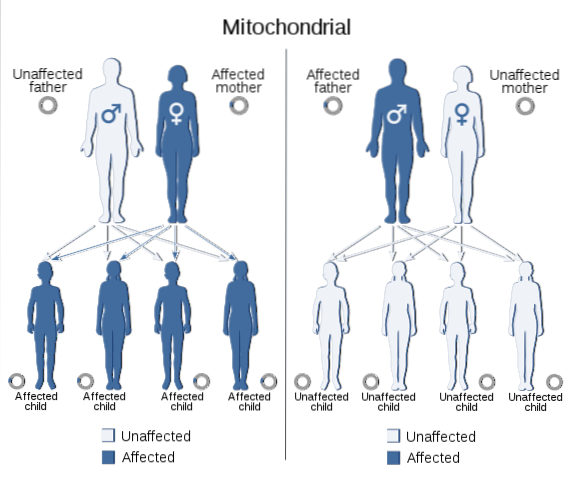
Quei tratti che derivano da mutazioni nel genoma mitocondriale mostrano uno specifico modello di segregazione che è stato chiamato "eredità mitocondriale", che di solito si verifica attraverso la linea materna, poiché l'ovulo fornisce il complemento totale del DNA mitocondriale e nessun mitocondrio è fornito dallo sperma.
L'imprinting genomico consiste in una serie di “segni” epigenetici che caratterizzano determinati geni o regioni genomiche complete e che risultano dal transito genomico del maschio o della femmina attraverso il processo di gametogenesi..
Esistono cluster di imprinting genico, che consistono tra 3 e 12 geni distribuiti tra 20 e 3700 chilogrammi di basi di DNA. Ogni cluster ha una regione nota come regione di controllo dell'imprinting, che mostra modifiche epigenetiche specifiche da ciascun genitore, tra cui:
- Metilazione del DNA su alleli specifici nei residui di citochine delle coppie CpG
- Modifica post-traduzionale degli istoni correlati alla cromatina (metilazione, acetilazione, fosforilazione, ecc., Delle code di amminoacidi di queste proteine).
Entrambi i tipi di “segni” modulano permanentemente l'espressione dei geni su cui si trovano, modificando i loro schemi di trasmissione alla generazione successiva..
I modelli di ereditarietà in cui l'espressione di una malattia dipende dagli alleli specifici ereditati da entrambi i genitori è noto come effetto di origine parentale..
Questo fenomeno è un'eccezione alla prima legge di Mendel, che afferma che solo uno dei due alleli presenti in ciascun genitore viene trasmesso alla prole e, secondo le leggi cromosomiche dell'ereditarietà, solo uno dei cromosomi omologhi parentali può essere trasmesso. la prossima generazione.
Questa è un'eccezione alla regola poiché la disomia uniparentale è l'eredità di entrambe le copie di un cromosoma omologa da uno dei genitori. Questo tipo di pattern ereditario non mostra sempre difetti fenotipici, poiché mantiene le caratteristiche numeriche e strutturali dei cromosomi diploidi..
Questo modello di ereditarietà consiste, dal punto di vista fenotipico, in una miscela di tratti codificati in allele che vengono combinati. Nei casi di dominanza incompleta, quegli individui che sono eterozigoti mostrano una miscela dei tratti dei due alleli che li controllano, il che implica che la relazione tra i fenotipi è modificata.
Descrive i modelli ereditari in cui i due alleli che vengono trasmessi dai genitori ai loro figli sono espressi simultaneamente in quelli con fenotipo eterozigote, per il quale entrambi sono considerati "dominanti".
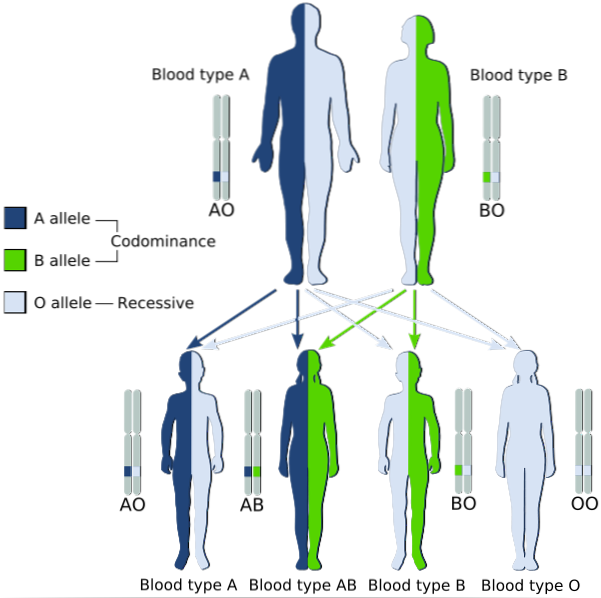
In altre parole, l'allele recessivo non è "mascherato" dall'espressione dell'allele dominante nella coppia allelica, ma entrambi sono espressi e nel fenotipo si osserva una miscela dei due tratti..
Forse uno dei principali punti deboli dell'eredità mendeliana è rappresentato dai tratti codificati da più di un allele, cosa abbastanza comune negli esseri umani e in molti altri esseri viventi..
Questo fenomeno ereditario aumenta la diversità dei tratti codificati da un gene e, inoltre, questi geni possono anche sperimentare modelli di dominanza incompleta e codominanza oltre alla dominanza semplice o completa..
Un'altra delle "pietre nella scarpa" o "gambe sciolte" delle teorie ereditarie di Mendel ha a che fare con quei geni che controllano la comparsa di più di un fenotipo o caratteristica visibile, come nel caso dei geni pleiotropici..
Nelle sue opere, Mendel inoltre non ha considerato l'eredità di alcuni alleli che possono impedire la sopravvivenza della prole quando è in forma omozigote o eterozigote; questi sono gli alleli letali.
Gli alleli letali sono solitamente correlati a mutazioni o difetti nei geni che sono strettamente necessari per la sopravvivenza, che quando trasmessi alla generazione successiva (tali mutazioni), a seconda dell'omozigosi o eterozigosi degli individui, sono letali.
Ci sono caratteristiche che sono controllate da più di un gene (con i loro alleli) e che, inoltre, sono fortemente controllate dall'ambiente. Negli esseri umani questo è estremamente comune ed è il caso di tratti come altezza, occhi, capelli e colore della pelle, nonché il rischio di soffrire di alcune malattie.
Nell'uomo e in molti animali ci sono anche tratti che si trovano su uno dei due cromosomi sessuali e che vengono trasmessi attraverso la riproduzione sessuale. Molti di questi tratti sono considerati "legati al sesso" quando sono evidenziati in uno solo dei sessi, sebbene entrambi siano fisicamente in grado di ereditare questi tratti..
La maggior parte dei tratti legati al sesso sono associati ad alcune malattie e disturbi recessivi.
Esiste una malattia genetica negli esseri umani nota come sindrome di Marfan, che è causata da una mutazione in un singolo gene che influenza simultaneamente la crescita e lo sviluppo (altezza, vista e funzione cardiaca, tra gli altri)..
Questo è un caso considerato un eccellente esempio del modello di ereditarietà non mendeliana chiamato pleiotropia, in cui un singolo gene controlla diverse caratteristiche.
Le malattie genetiche che derivano da mutazioni nel DNA mitocondriale presentano una serie di variazioni fenotipiche cliniche poiché si verifica ciò che è noto come eteroplasmia, dove diversi tessuti hanno una diversa percentuale di genoma mitocondriale mutante e, quindi, presentano diversi fenotipi.
Tra questi disturbi ci sono le sindromi da "deplezione" mitocondriale, che sono un gruppo di malattie autosomiche recessive caratterizzate da una significativa riduzione del contenuto di DNA mitocondriale, che si conclude con sistemi di produzione di energia carenti in quegli organi e tessuti più colpiti..
Queste sindromi possono essere dovute a mutazioni nel genoma nucleare che interessano i geni nucleari coinvolti nella sintesi dei nucleotidi mitocondriali o nella replicazione del DNA mitocondriale. Gli effetti possono essere evidenziati come miopatie, encefalopatie, difetti epato-cerebrali o neuro-gastrointestinali.


Nessun utente ha ancora commentato questo articolo.