Il eritropoiesi È il processo attraverso il quale si formano i globuli rossi o gli eritrociti. Questi globuli, nell'uomo, hanno una vita media di 4 mesi e non sono in grado di riprodursi. A causa di ciò, è necessario creare nuovi eritrociti per sostituire quelli che muoiono o si perdono nelle emorragie..
Negli uomini, il numero di globuli rossi è di circa 54 milioni per millilitro, mentre nelle donne è leggermente inferiore (48 milioni). Ogni giorno vengono persi circa 10 milioni di eritrociti, quindi una quantità simile deve essere sostituita.
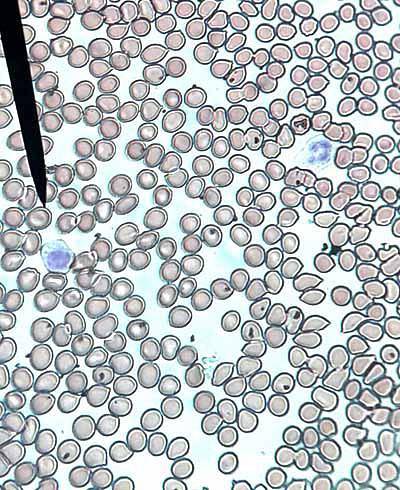
Gli eritrociti sono formati da eritroblasti nucleati presenti nel midollo osseo rosso dei mammiferi, mentre in altri vertebrati sono prodotti principalmente nei reni e nella milza.
Quando raggiungono la fine dei loro giorni, si frammentano; quindi le cellule chiamate macrofagi li circondano. Questi macrofagi sono presenti nel fegato, nel midollo osseo rosso e nella milza..
Quando i globuli rossi vengono distrutti, il ferro viene riciclato per essere riutilizzato, mentre il resto dell'emoglobina si trasforma in un pigmento biliare chiamato bilirubina..
L'eritropoiesi è stimolata da un ormone chiamato eritropoietina, ma il processo è regolato da diversi fattori, come la temperatura, la pressione dell'ossigeno, tra gli altri..
Indice articolo
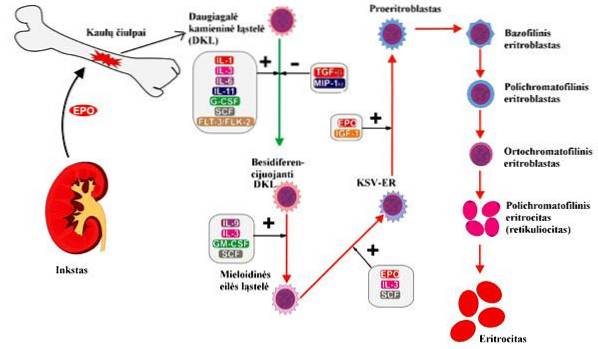
Negli organismi adulti, l'eritropoiesi si verifica in siti specializzati nel midollo osseo rosso chiamati isole eritroblastiche. Per la formazione degli eritrociti devono avvenire diversi processi, che vanno dalla proliferazione cellulare alla maturazione dei globuli rossi, passando per vari stadi di differenziazione cellulare..
Quando le cellule subiscono divisioni mitotiche, la loro dimensione e quella del loro nucleo diminuiscono, così come la condensazione della cromatina e l'emoglobinizzazione. Inoltre si stanno allontanando dalla zona di origine.
Nelle fasi finali, perderanno il nucleo e altri organelli ed entreranno in circolazione, migrando attraverso i pori citoplasmatici delle cellule endoteliali..
Alcuni autori dividono l'intero processo di eritropoiesi in due fasi, la prima di proliferazione e differenziazione cellulare; mentre altri dividono il processo in base alle caratteristiche specifiche della cellula in ogni fase, quando osservati con la macchia di Wright. Sulla base di quest'ultimo, le fasi dell'eritropoiesi sono:
Sono le prime cellule sensibili all'eritropoietina, alcuni autori le chiamano progenitori mieloidi, o anche BFU-E, per il suo acronimo in inglese. Sono caratterizzati dall'espressione di antigeni di superficie come il CD34, nonché dalla presenza di recettori dell'eritropoietina in piccole quantità.
Abbreviati in inglese come CFU-E, sono in grado di produrre piccole colonie di eritroblasti. Un'altra caratteristica di queste cellule è che le quantità di recettori dell'eritropoietina sono molto più elevate rispetto alle unità formanti colonie burst..
Considerato come il primo stadio di maturazione degli eritrociti. Si caratterizzano per le grandi dimensioni (da 14 a 19 µm secondo alcuni autori, fino a 25 µm secondo altri). Il nucleo è arrotondato e presenta anche nucleoli e abbondante cromatina.
Considerato come il primo stadio di maturazione degli eritrociti. Si caratterizzano per le grandi dimensioni (da 14 a 19 µm secondo alcuni autori, fino a 25 µm secondo altri). Il nucleo è grande, arrotondato, con la cromatina disposta sotto forma di filamenti e 2 o 3 nucleoli.
L'assorbimento del ferro plasmatico inizia in questa fase. Hanno un'emivita di 20 ore, per lasciare il posto alla fase successiva attraverso la mitosi.
Chiamati anche normoblasti, sono più piccoli dei loro precursori. Queste cellule si colorano di blu con una colorazione vitale, cioè sono basofile. Il nucleo è condensato, i nucleoli sono scomparsi e hanno un gran numero di ribosomi. In questa fase inizia la sintesi dell'emoglobina.
All'inizio sono conosciuti come eritroblasti basofili di Tipo I e dopo una divisione mitotica si trasformano in Tipo II, che rimangono basofili e presentano una maggiore sintesi di emoglobina. La durata approssimativa di entrambe le cellule, insieme, è simile a quella dei proeritroblasti.
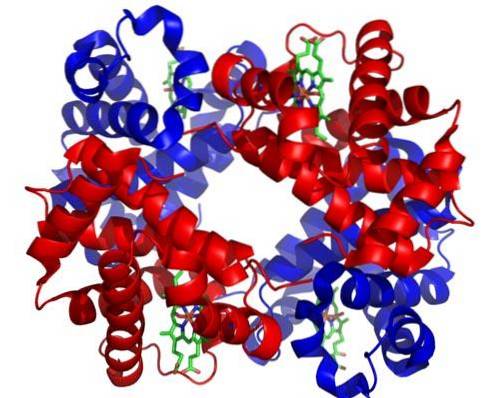
Sono formati dalla divisione mitotica degli eritroblasti basofili di tipo II e sono le ultime cellule in grado di dividersi per mitosi. Le loro dimensioni variano da 8 a 12 µm e hanno un nucleo arrotondato e condensato.
Il citoplasma di queste cellule si colora di grigio piombo con la colorazione di Wright. Ha un'alta concentrazione di emoglobina e il numero di ribosomi rimane elevato.
6-eritroblasti ortocromatici
Il colore di queste cellule è rosa o rosso a causa della quantità di emoglobina che hanno. La sua dimensione è leggermente inferiore a quella dei suoi precursori (da 7 a 10 µm) e ha un piccolo nucleo, che verrà espulso dall'esocitosi quando le cellule matureranno..
Sono formati dalla differenziazione degli eritroblasti ortocromici, perdono gli organelli e riempiono il loro citoplasma di emoglobina. Rimangono nel midollo osseo rosso per due o tre giorni fino a quando non migrano nel sangue dove completeranno la loro maturazione..
Sono gli elementi formati maturi, il prodotto finale dell'eritropoiesi e che sono formati dalla maturazione dei reticolociti. Hanno una forma biconcava per l'assenza di un nucleo e per l'interazione tra il citoscheletro eritrocitario e due proteine chiamate spettrina e actina.
Sono i globuli più abbondanti, sono formati da reticolociti. Nei mammiferi hanno una forma biconcava a causa dell'assenza di un nucleo e dell'interazione tra il citoscheletro eritrocitario e due proteine chiamate spettrina e actina. In altri vertebrati sono arrotondati e trattengono il nucleo.
Sebbene l'eritropoietina stimoli la formazione dei globuli rossi per migliorare la capacità di trasporto dell'ossigeno del sangue, esistono diversi meccanismi fondamentali per regolare questa formazione, tra cui:
La concentrazione di ossigeno nel sangue regola l'eritropoiesi. Quando questa concentrazione è molto bassa nel flusso sanguigno al rene, viene stimolata la produzione di globuli rossi..
Questa bassa concentrazione di O2 tissutale può verificarsi a causa di ipossiemia, anemia, ischemia renale o quando l'affinità dell'emoglobina per questo gas è superiore al normale..
Miescher, nel 1893, fu il primo a suggerire la relazione tra ipossia tissutale ed eritropoiesi. Tuttavia, questa ipossia non stimola direttamente il midollo osseo a produrre globuli rossi, come suggerito da Miescher. Al contrario, induce il rene a produrre l'ormone eritropoietina.
La produzione di eritropoietina dovuta all'ipossia tissutale è regolata geneticamente ei recettori che rilevano tale ipossia si trovano all'interno del rene. Anche la produzione di eritropoietina è aumentata a causa di un calo della pressione parziale dell'ossigeno nei tessuti dopo il sanguinamento..
Le cellule che producono l'eritropoietina si trovano nei reni e nel fegato. L'aumento della produzione di questo ormone durante l'anemia è dovuto ad un aumento del numero di cellule che lo producono..
Il testosterone regola indirettamente l'eritropoiesi regolando i livelli di ferro nel sangue. Questo ormone agisce direttamente sull'azione di una proteina citoplasmatica chiamata BMP-Smad (proteina morfogenetica ossea-Smad) negli epatociti.
A causa dell'azione del testosterone, la trascrizione dell'epcidina viene soppressa. Questa epcidina impedisce il passaggio del ferro dalle cellule nel plasma dai macrofagi che riciclano il ferro, portando a una drastica diminuzione del ferro nel sangue.
Quando si verifica l'ipoferremia, ci sarà un'inibizione dell'eritropoietina, poiché non ci sarà ferro per la produzione di eritrociti.
È stato dimostrato che la temperatura ha un effetto sull'eritripoiesi. L'esposizione a temperature molto basse causa la necessità di produrre calore nei tessuti.
Ciò richiede l'aumento del numero di eritrociti per fornire ossigeno ai tessuti periferici. Tuttavia, non è del tutto chiaro come avvenga questo tipo di regolazione..
A quanto pare, c'è una produzione di eritropoietina da parte dei neuroni del sistema nervoso centrale, per proteggersi dal danno ischemico e dall'apoptosi. Tuttavia, gli scienziati non sono stati ancora in grado di verificarlo..
Gli agenti stimolanti l'eritropoiesi (ESA) sono agenti responsabili della stimolazione della produzione di eritrociti. L'eritropoietina è l'ormone naturalmente responsabile di questo processo, ma esistono anche prodotti sintetici con proprietà simili.
L'eritropoietina è un ormone sintetizzato principalmente a livello renale. Durante le prime fasi dello sviluppo, il fegato è anche coinvolto nella produzione attiva di eritropoietina. Tuttavia, man mano che lo sviluppo procede, quest'ultimo organismo è meno coinvolto nel processo..
L'eritrocita inizia a disperdere i recettori per l'eritropoietina sulla superficie della membrana. L'eritropoietina attiva una serie di cascate di trasduzione del segnale intercellulare che inizialmente producono la sintesi dell'emoglobina e fanno sì che i reticolociti agiscano più rapidamente e vengano rilasciati nella circolazione..
Gli ESA artificiali sono classificati in generazioni (dalla prima alla terza), a seconda della data in cui sono stati creati e commercializzati. Strutturalmente e funzionalmente simile all'eritropoietina.
Gli ESA di prima generazione sono noti come epoetina alfa, beta e delta. I primi due sono prodotti per ricombinazione da cellule animali e hanno un'emivita di circa 8 ore nel corpo. L'epoetina delta, nel frattempo, viene sintetizzata dalle cellule umane.
Darbepoetin alfa è un ESA di seconda generazione, prodotto da cellule di criceto cinese utilizzando una tecnologia chiamata DNA ricombinante. Ha un'emivita tre volte superiore a quella degli ESA di prima generazione. Come con le epoetine, alcuni atleti ad alte prestazioni hanno utilizzato la darbepoetina come mezzo di doping.
Continuous Erythropoetin Receptor Activator, o CERA per il suo acronimo in inglese, è il nome generico degli ESA di terza generazione. Non tentano di simulare la struttura e la funzione dell'eritropoietina, ma agiscono piuttosto stimolando il recettore dell'eritropoietina, aumentandone così gli effetti..
La sua emivita è di diverse settimane invece di ore, come i farmaci precedenti. Utilizzato commercialmente dal 2008, tuttavia, il suo uso illecito in attività sportive risale apparentemente a due o tre anni prima della sua commercializzazione legale.
L'eritropoiesi inefficace o inefficace si verifica quando i globuli rossi formati sono difettosi e di solito vengono distrutti prima o subito dopo aver lasciato il midollo osseo..
L'eritropoiesi inefficace può essere dovuta a difetti nella sintesi degli acidi nucleici, del gruppo eme o delle globine. Questi difetti causano diversi tipi di anemia.
In questo caso, c'è una carenza di acido folico e cobalamina, la sintesi del DNA è inibita nel nucleo delle cellule promotrici degli eritrociti, in modo che non siano in grado di dividersi mitoticamente. Il citoplasma, da parte sua, aumenta il suo volume (macrocitosi), dando origine a una grande cellula chiamata megaloblasto.
In questi casi si originano una serie di anemie chiamate anemie megaloblastiche, di cui la più comune è l'anemia perniciosa. In questa malattia non c'è assorbimento della vitamina B12 nell'intestino tenue.
Altre cause di anemia megaloblastica includono malattie digestive, malassorbimento, carenza di acido folico e a causa di alcuni farmaci.
I sintomi di questo tipo di anemia includono pallore anormale, irritabilità, perdita di appetito, diarrea, difficoltà a camminare o debolezza muscolare. A seconda della causa, può essere trattata con integratori vitaminici o di acido folico..
L'eritropoiesi inefficace a causa della carenza nella sintesi del ferro può causare due tipi di anemia; anemia microcitica da carenza di ferro e anemia sideroblastica.
L'anemia microcitica è nota come un gruppo di anemie caratterizzate da globuli rossi piccoli e pallidi, possono avere origini diverse, tra cui la talassemia e l'eritropoiesi inefficace.
I livelli di ferro ed emosiderina sono molto alti nell'anemia sideroblastica. L'emosiderina è un pigmento giallo derivato dall'emoglobina e compare quando i livelli del metallo sono più alti del normale. Questo tipo di anemia provoca la morte dei basofili nel midollo osseo rosso e non c'è sintesi di emoglobina.
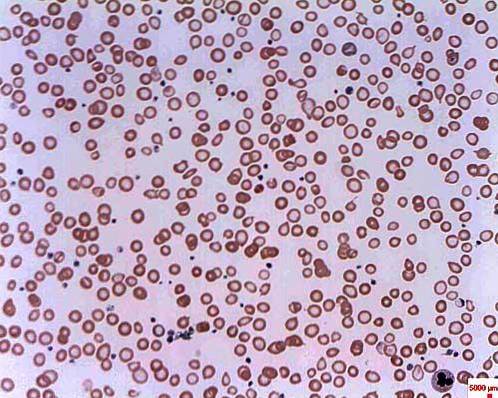
Si chiama anemia sideroblastica perché gli eritrociti si sviluppano in modo anomalo a causa dell'accumulo di ferro sotto forma di granuli, ricevendo il nome di sideroblasti. L'anemia sideroblastica può essere congenita o secondaria e avere cause diverse.
In questo caso, si verificano anemia falciforme e beta talassemia. L'anemia falciforme è anche nota come anemia falciforme. È prodotto da una mutazione genetica che porta alla sostituzione dell'acido glutammico con la valina durante la sintesi della beta globina.
A causa di questa sostituzione, l'affinità dell'emoglobina per l'ossigeno diminuisce e si verifica un'atrofia dell'eritrocita, che assume una forma a falce invece della normale forma a disco biconcava. Il paziente con anemia falciforme è suscettibile a microinfarti ed emolisi.
La talassemia è una malattia causata da una codifica genetica inadeguata delle globine α e β che porta a una morte precoce dell'eritrocita. Esistono circa un centinaio di mutazioni diverse che possono causare talassemia con diversi gradi di gravità.


Nessun utente ha ancora commentato questo articolo.