Il uracile È una base azotata di tipo pirimidinico, che si trova nell'acido ribonucleico (RNA). Questa è una delle caratteristiche che differenziano l'RNA dall'acido desossiribonucleico (DNA), poiché quest'ultimo ha timina al posto dell'uracile. Entrambe le sostanze, uracile e timina, differiscono solo per il fatto che la seconda ha un gruppo metilico.
Da un punto di vista evolutivo, è stato proposto che l'RNA fosse la prima molecola a immagazzinare informazioni genetiche e funzionare come catalizzatore nelle cellule, prima del DNA e degli enzimi. Per questo motivo, si ritiene che l'uracile abbia svolto un ruolo chiave nell'evoluzione della vita.
Negli esseri viventi, l'uracile non si trova in una forma libera, ma comunemente forma nucleotidi monofosfato (UMP), difosfato (UDP) e trifosfato (UTP). Questi nucleotidi uracilici hanno funzioni diverse, come la biosintesi dell'RNA e del glicogeno, l'interconversione isomerica degli zuccheri e la regolazione della glutammina sintasi..
Indice articolo
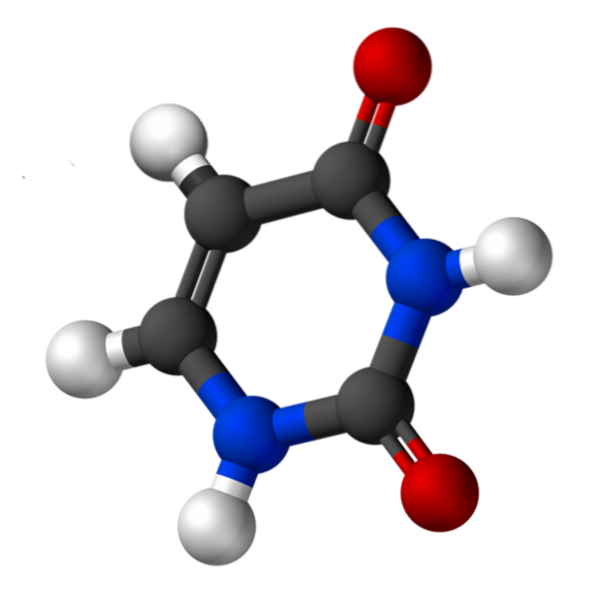
L'uracile, chiamato 2,4-diossipiridina, ha la formula empirica C4H4NDueODue, il cui peso molecolare è 112,09 g / mol, e viene purificato come polvere bianca.
La struttura dell'uridina è un anello eterociclico con quattro atomi di carbonio e due atomi di azoto, con doppi legami alternati. È planare.
Ha una solubilità di 50 mg / ml, a 25ºC, in idrossido di sodio 1M e un pKa compreso tra 7,9 e 8,2. La lunghezza d'onda in cui si verifica la sua massima assorbanza (ʎmax) è compreso tra 258 e 260 nm.
Esiste una via comune per la biosintesi dei nucleotidi pirimidinici (uracile e citochina). Il primo passo è la biosintesi del carbamoil fosfato da CODue e NH4+, che è catalizzato dalla carbamoil fosfato sintetasi.
La pirimidina è costituita da carboil fosfato e aspartato. Entrambe le sostanze reagiscono e formano N-carbamoilaspartato, una reazione catalizzata dall'aspartato transcabamoilasi (ATCasi). La chiusura dell'anello pirimidinico è causata dalla disidratazione catalizzata dalla diidrootasi e produce L-diidrorotato..
L'L-diidrorotato viene ossidato e convertito in orotato; l'accettore di elettroni è NAD+. È una reazione catalizzata dalla diidroorotato deidrogenasi. Il passaggio successivo è il trasferimento del gruppo fosforibosile, dal pirofosfato fosforibosilico (PRPP), all'orotato. Forma orotidilato (OMP) e pirofosfato inorganico (PPi), catalizzato da orotato fosforibosil transferasi.
L'ultimo passaggio consiste nella decarbossilazione dell'anello pirimidinico dell'orotidilato (OMP). Forma uridilato (uridina-5'-monofosfato, UMP), che è catalizzato da una decarbossilasi.
Quindi, attraverso la partecipazione di una chinasi, un gruppo fosfato viene trasferito da ATP a UMP, formando UDP (uridina-5'-difosfato). Quest'ultimo si ripete, formando UTP (uridina-5'-trifosfato).
Nei batteri, la regolazione della biosintesi della pirimidina avviene tramite feedback negativo, a livello dell'aspartato transcabamoilasi (ATCasi).
Questo enzima è inibito dal CTP (citidina-5'-trifosfato), che è il prodotto finale della via biosintetica della pirimidina. ATCase ha subunità regolatorie che si legano al regolatore allosterico CTP.
Negli animali la regolazione della biosintesi della pirimidina avviene tramite feedback negativo, a livello di due enzimi: 1) carbamoil fosfato sintasi II, inibito dall'UTP e attivato da ATP e PRPP; e 2) OMP decarbossilasi, che è inibita dal prodotto della reazione che catalizza, UMP. Il tasso di biosintesi di OMP varia con la disponibilità di PRPP.
L'uracile è presente in tutti i tipi di RNA, come l'RNA messaggero (mRNA), l'RNA di trasferimento (tRNA) e l'RNA ribosomiale (rRNA). La biosintesi di queste molecole avviene attraverso un processo chiamato trascrizione..
Durante la trascrizione, le informazioni contenute nel DNA vengono copiate nell'RNA da una RNA polimerasi. Il processo inverso, in cui le informazioni contenute nell'RNA vengono copiate nel DNA, si verifica in alcuni virus e piante attraverso la trascrittasi inversa..
La biosintesi dell'RNA richiede nucleoside trifosfato (NTP), vale a dire: uridina trifosfato (UTP), citidina trifosfato (CTP), adenina trifosfato (ATP) e guanina trifosfato (GTP). La reazione è:
(RNA)n residui + NTP -> (RNA)n + 1 residuo + PPi
L'idrolisi del pirofosfato inorganico (PPi) fornisce l'energia per la biosintesi dell'RNA.
Gli esteri dello zucchero sono molto comuni negli organismi viventi. Alcuni di questi esteri sono difosfati di esteri nucleosidici, come gli zuccheri UDP, che sono molto abbondanti nelle cellule. Gli zuccheri UDP partecipano alla biosintesi di disaccaridi, oligosaccaridi e polisaccaridi.
Nelle piante, la biosintesi del saccarosio avviene attraverso due vie: una primaria e una secondaria.
La via principale è il trasferimento del D-glucosio da UDP-D-glucosio a D-fruttosio per formare saccarosio e UDP. La via secondaria comprende due fasi: inizia con UDP-D-glucosio e fruttosio-6-fosfato e termina con la formazione di saccarosio e fosfato..
Nelle ghiandole mammarie, la biosintesi del lattosio si verifica da UDP-D-galattosio e glucosio.
Nelle piante, la biosintesi della cellulosa viene effettuata dalla condensazione continua di residui beta-D-glucosilici, dall'UDP-glucosio all'estremità non riducente della catena del poliglucosio in crescita. Allo stesso modo, la biosintesi di amilosio e amilopectina richiede UDP-glucosio come substrato donatore di glucosio per la catena in crescita..
Negli animali, sia l'UDP-glucosio che l'ADP-glucosio vengono utilizzati per la biosintesi del glicogeno. Allo stesso modo, la biosintesi del condroitin solfato richiede UDP-xilosio, UDP-galattosio e UDP-glucuronato..
La conversione del galattosio in un intermedio di glicolisi avviene attraverso il percorso di Leloir. Uno dei passaggi di questo percorso è catalizzato dall'enzima UDP-galattosio-4-epimerasi, che facilita l'interconversione dell'UDP-galattosio in UDP-glucosio..
Durante la biosintesi della glicoproteina, le proteine attraversano i sacchi cis, medio e trans dell'apparato di Golgi.
Ciascuna di queste sacche ha una serie di enzimi che processano le glicoproteine. I monomeri dello zucchero, come il glucosio e il galattosio, vengono aggiunti all'oligosaccaride proteico da UDP-esosio e altri nucleotidi-esosio.
I nucleotidi esosi vengono trasportati alle cisterne del Golgi da antiport. L'UDP-galattosio (UDP-Gal) e l'UDP-N-acetilgalattosamina (UDP-GalNAc) entrano nelle cisterne dal citosol scambiandosi con l'UMP.
Nella cisterna Golgi, una fosfatasi idrolizza un gruppo fosfato su UDP e forma UMP e Pi. L'UDP proviene da reazioni catalizzate dalla galattosiltransferasi e dalla N-acetilgalattosamiltransferasi. L'UMP formato dalla fosfatasi serve per lo scambio nucleotide-esoso.
Un meccanismo di regolazione della glutammina sintasi è la modificazione covalente, che consiste nell'adenilazione, che la inattiva, e nella dedenilazione, che la attiva. Questa modifica covalente è reversibile e catalizzata dall'adeniltransferasi..
L'attività dell'adeniltransferasi è modulata dal legame della proteina PII, che è regolata da una modifica covalente, l'uridinilazione.
Sia l'uridilazione che la deuridilazione vengono eseguite dall'uridililtransferasi. In questo enzima, l'attività di uridilazione è dovuta alla glutammina e al fosfato e viene attivata dal legame di alfa-chetoglutarato e ATP ai PII.
Alcuni mRNA vengono modificati prima della traduzione. In alcuni organismi eucarioti, come Trypanosoma brucei, c'è un editing dell'RNA del trascritto del gene della subunità II della citocromo ossidasi. Ciò avviene attraverso l'inserimento di residui di uracile, una reazione catalizzata dall'uridiltransferasi terminale..
Un RNA guida, complementare al prodotto modificato, funge da modello per il processo di modifica. Le coppie di basi formate tra la trascrizione iniziale e l'RNA guida implicano coppie di basi G = U che non sono Watson-Crick e sono comuni nell'RNA..
In condizioni fisiologiche, la biosintesi del glicogeno dal glucosio-1-fosfato è termodinamicamente impossibile (ΔG positivo). Per questo motivo, prima della biosintesi, si verifica l'attivazione del glucosio-1-fosfato (G1P). Questa reazione combina G1P e UTP per formare glucosio uridina difosfato (UDP-glucosio o UDPG)..
La reazione è catalizzata dalla pirofosforilasi UDP-glucosio ed è la seguente:
G1P + UTP -> UDP-glucosio + 2Pi.
La variazione di energia libera di Gibbs in questo passaggio è ampia e negativa (-33,5 KJ / mol). Durante la reazione all'ossigeno, G1P attacca l'atomo di fosforo alfa di UTP e forma UDP-glucosio e pirofosfato inorganico (PPi). Successivamente, il PPi viene idrolizzato da una pirofosfatasi inorganica, la cui energia di idrolisi è ciò che guida la reazione generale.
L'UDP-glucosio è una sostanza "ad alta energia". Permette di formare i legami glicosidici tra il residuo di glucosio e la catena polisaccaridica in crescita. Questo stesso principio energetico è applicabile alle reazioni in cui partecipano gli zuccheri UDP, come la biosintesi di disaccaridi, oligosaccaridi e glicoproteine..
Ci sono lesioni al DNA che si verificano spontaneamente. Una di queste lesioni è la deaminazione spontanea delle citochine e la sua conseguente conversione in uracile. In questo caso, la riparazione avviene rimuovendo la base di DNA modificata da un enzima chiamato uracile DNA glicosilasi..
L'enzima uracile DNA glicosilasi rimuove la citochina danneggiata (uracile), producendo un residuo di desossiribosio privo della base azotata, chiamato sito AP (sito apurinico-apirimidinico).
L'enzima AP endonucleasi taglia quindi la spina dorsale del fosfodiestere del sito AP, rimuovendo il residuo zucchero-fosfato. La DNA polimerasi I ripristina il filamento danneggiato.



Nessun utente ha ancora commentato questo articolo.