
Il recettori di membrana Sono un tipo di recettori cellulari che si trovano sulla superficie della membrana plasmatica delle cellule, consentendo loro di rilevare sostanze chimiche che per loro natura non possono attraversare la membrana.
In generale, i recettori di membrana sono proteine di membrana integrali specializzate nella rilevazione di segnali chimici come ormoni peptidici, neurotrasmettitori e alcuni fattori trofici; anche alcuni farmaci e tossine possono legarsi a questi tipi di recettori.
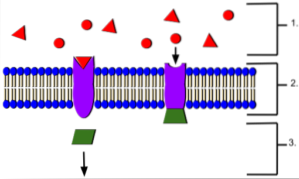
Sono classificati in base al tipo di cascata intracellulare a cui sono accoppiati e quali sono quelli che determinano l'effetto finale sulla cellula corrispondente, chiamata cellula bersaglio o cellula bersaglio..
Pertanto, sono stati descritti tre grandi gruppi: quelli legati ai canali ionici, quelli legati agli enzimi e quelli legati alla proteina G. Il legame dei ligandi ai recettori genera un cambiamento conformazionale nel recettore che innesca una cascata di segnali intracellulari nella cellula bersaglio.
Le catene di segnalazione accoppiate ai recettori di membrana consentono di amplificare i segnali e generare risposte o cambiamenti transitori o permanenti nella cellula bersaglio. Questi segnali intracellulari sono chiamati collettivamente "sistema di trasduzione del segnale"..
Indice articolo
La funzione dei recettori di membrana, e di altri tipi di recettori in generale, è quella di consentire la comunicazione delle cellule tra loro, in modo tale che i diversi organi e sistemi di un organismo lavorino in modo coordinato per mantenere l'omeostasi e rispondere a comandi volontari e automatici emessi dal sistema nervoso.
Pertanto, un segnale chimico che agisce sulla membrana plasmatica può innescare una modifica amplificata di varie funzioni all'interno del meccanismo biochimico di una cellula e innescare una molteplicità di risposte specifiche.
Attraverso il sistema di amplificazione del segnale, un singolo stimolo (ligando) è in grado di generare cambiamenti transitori immediati e indiretti e cambiamenti a lungo termine, modificando l'espressione di alcuni geni all'interno della cellula bersaglio, ad esempio.
I recettori cellulari sono classificati, in base alla loro posizione, in: recettori di membrana (quelli che sono esposti nella membrana cellulare) e recettori intracellulari (che possono essere citoplasmatici o nucleari).
I recettori di membrana sono di tre tipi:
- Legato a canali ionici
- Legato agli enzimi
- Legato alla proteina G.
Chiamati anche canali ionici ligando-gated, sono proteine di membrana composte da 4 a 6 subunità che sono assemblate in modo tale da lasciare un canale centrale o poro, attraverso il quale gli ioni passano da un lato della membrana.
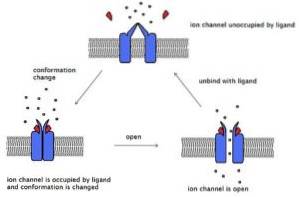
Questi canali attraversano la membrana e hanno un'estremità extracellulare, dove si trova il sito di legame per il ligando, e un'altra estremità intracellulare che, in alcuni canali, ha un meccanismo di gate. Alcuni canali hanno un sito ligando intracellulare.
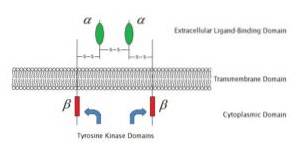
Questi recettori sono anche proteine transmembrana. Hanno un'estremità extracellulare che presenta il sito di legame per il ligando e che hanno associato alla loro estremità intracellulare un enzima che viene attivato dal legame del ligando al recettore.
I recettori accoppiati a proteine G hanno un meccanismo indiretto per la regolazione delle funzioni intracellulari delle cellule bersaglio che coinvolge le molecole del trasduttore chiamate proteine leganti o GTP o proteine G..
Tutti questi recettori legati alla proteina G sono costituiti da una proteina di membrana che attraversa la membrana sette volte e sono chiamati recettori metabotropici. Sono stati identificati centinaia di recettori legati a diverse proteine G..
Nei recettori legati ai canali ionici, il legame del ligando al recettore genera un cambiamento conformazionale nella struttura del recettore che può modificare un gate, avvicinare o allontanare le pareti del canale. Ciò modifica il passaggio degli ioni da un lato all'altro della membrana..
I recettori legati ai canali ionici sono, per la maggior parte, specifici per un tipo di ione, motivo per cui sono stati descritti i recettori per i canali K +, Cl-, Na +, Ca ++, ecc. Esistono anche canali che lasciano passare due o più tipi di ioni.
La maggior parte dei recettori legati agli enzimi si associa alle protein chinasi, in particolare all'enzima tirosin chinasi. Queste chinasi vengono attivate quando il ligando si lega al recettore nel suo sito di legame extracellulare. Le chinasi fosforilano proteine specifiche nella cellula bersaglio, modificandone la funzione.
I recettori legati alle proteine G attivano cascate di reazioni biochimiche che finiscono per modificare la funzione di varie proteine nella cellula bersaglio..
Esistono due tipi di proteine G che sono le proteine G eterotrimeriche e le proteine G monomeriche. Entrambi sono legati al PIL in modo inattivo, ma quando il ligando è legato al recettore, il PIL viene sostituito da GTP e la proteina G viene attivata..
Nelle proteine G eterotrimeriche, la subunità α legata a GTP si dissocia dal complesso ßγ, lasciando la proteina G attivata. Sia la subunità α legata a GTP che la ßγ libera possono mediare la risposta.

Le proteine monomeriche G o piccole proteine G sono anche chiamate proteine Ras perché sono state descritte per la prima volta in un virus che produce tumori sarcomatosi nei ratti..
Quando attivati, stimolano meccanismi legati principalmente al traffico vescicolare e alle funzioni citoscheletriche (modifica, rimodellamento, trasporto, ecc.).
Il recettore dell'acetilcolina, collegato a un canale del sodio che si apre al legame con l'acetilcolina e provoca la depolarizzazione della cellula bersaglio, è un buon esempio di recettori di membrana legati ai canali ionici. Inoltre, ci sono tre tipi di recettori del glutammato che sono recettori ionotropici..
Il glutammato è uno dei più importanti neurotrasmettitori eccitatori del sistema nervoso. I suoi tre tipi di recettori ionotropici sono: recettori NMDA (N-metil-D-aspartato), AMPA (α-ammino-3-idrossi-5-metil-4-isossazolo-propionato) e kainato (acido kainico).
I loro nomi derivano dagli agonisti che li attivano e questi tre tipi di canali sono esempi di canali eccitatori non selettivi, poiché consentono il passaggio di sodio e potassio e in alcuni casi piccole quantità di calcio..
Esempi di recettori legati agli enzimi sono il recettore dell'insulina, la famiglia di recettori TrK o recettori neurotrofine e i recettori per alcuni fattori di crescita..
I principali recettori accoppiati a proteine G includono recettori muscarinici dell'acetilcolina, recettori β-adrenergici, recettori del sistema olfattivo, recettori metabotropici del glutammato, recettori per molti ormoni peptidici e recettori della rodopsina del sistema retinico..


Nessun utente ha ancora commentato questo articolo.